









Scrivo questo articolo a seguito delle tante domande che mi vengono poste sul forum, come questa: se accoppio quella forma con quell’altra che esce fuori?
La genetica dei Betta è un vero rompicapo, spesso vedo sul web espertoni che se ne escono con strane tavole di Mendel , ma la genetica dei Betta non ha una regola precisa essendo frutto di lunghe selezioni spesso incrociando specie affini. Si può parlare di probabilità ma no di risultati sicuri quando si dice che nasceranno di quel colore o di quella forma, la genetica dei Betta va oltre le famose tavole di Mendel, che riporterò anche in questo articolo, ma che come dirò vanno prese con le pinze per i motivi che elencherò di seguito, quella è la parte teorica, la pratica è ben diversa.
Se la genetica delle forme ha le sue difficoltà ancora più complessa è quella delle colorazioni, li c’è veramente da uscirne matti.
Attenzione la genetica nelle forme ha il suo ruolo fondamentale ma ha il suo ruolo anche la qualità dell’acqua in cui i Betta crescono e la loro alimentazione, come ho più volte detto sul forum. Personalmente riesco spesso a capire se un Betta è nato in Europa o in Asia , perchè ho riscontrato spesso li stessi difetti in moltissimi Betta nati in Europa, so che questo concetto può star scomodo a qualcuno, ma per me è la verità come più volte ho provato. Infatti, spesso per via dell’acqua, accoppiando anche dei bellissimi Halfmoon Asiatici , nelle discendenze si perdono certe caratteristiche, un pò perchè ritengo gli allevatori europei di certo non a livello di quelli Asiatici ma più scarsi, ma anche per il fattore acqua. A volte utenti mi hanno fatto vedere Betta nati da HM europei , poi riprodotti da loro, e presentavano la coda arrotondata, attenzione quello non è un difetto dovuto all’acqua come spesso si crede ma dovuto in maggioranza alla genetica, l’allevamento nelle acque europee da problemi nella crescita delle ramificazioni, quindi spesso abbiamo come carattere fenotipico, cioè non dovuto alla genetica ma da fattori esterni, pinne mal ramificate, poco rigide con iper ramificazioni tipo rosetail o feathertail , attenzione il fattore rosetail genetico è diverso da quello fenotipico, sono 2 difetti simili uno genetico e uno fenotipico per via delle nostre acque.
Quindi per prima cosa, chiudendo questa introduzione, sognatevi di ottenere Betta Show come quelli asiatici, e se qualcuno vi dice che alleva Betta della stessa qualità di quelli Asiatici sta a voi crederci o no. Come sempre invito sempre tutti a consultarci sul forum prima di comprare qualsiasi Betta sia se Europeo che Asiatico. Faccio questo discorso perchè lo ritengo importante , per me è anche controproducente essendo allevatore, però ci tengo a farlo viste le mille fregature prese da miei amici neofiti spendendo troppi soldi su Betta Europei. Non sto dicendo che tutti i Betta Europei fanno schifo, ma solo che per me, e a mia esperienza, quelli Europei sono sempre un gradino sotto come qualità selettiva a quelli Asiatici.
Veniamo all’argomento base di questo articolo, la genetica delle forme, la genetica ci dice cosa esce fuori se accoppiamo quella forma con quell’altra , dandoci anche le percentuali delle varie forme che possono uscire. Parliamo di percentuali e probabilità perchè in primis dipende da quante uova vengono fecondate e quali, se per esempio avviene la fecondazione dell’unico uovo che presenta tutti i caratteri recessivi , noi avremo solo un piccolo con i caratteri recessivi,cioè quello che aveva meno probabilità di uscire, ma essendo solo lui rappresenterebbe la maggioranza . Altro fattore importante, per poter dire con precisione cosa nascerà da una coppia bisognerebbe conoscere almeno in parte la loro discendenza , per capire che caratteri portano con se ma che non manifestano , esempio: Accoppiamo 2 HM , ma uno è un’ HM puro di almeno 3 generazioni, l’altro invece è figlio di un accoppiamento fra SD e HM , ed essendo il carattere SD dominante darà solo pochissimi HM reali , noi non sapendolo accoppiamo questi 2 HM e crediamo, che come normalmente avviene, nasceranno in maggioranza HM , in questo caso invece si ripresenterà il carattere SD e i veri HM saranno pochissimi. Quindi per prima cosa quando si vuole selezionare una forma comprate forme pure (meglio se Asiatiche), e accoppiate le coppie più volte per poter avere più esemplari con i caratteri che vogliamo.
Io non spiegherò la genetica delle forme solo con i classici schemi di Mendel, perchè so che spesso a chi è neofita sono di difficile comprensione, riporterò come mio solito in primis esempi pratici.
Per prima cosa vediamo come si definiscono le generazioni, se prendiamo una coppia e la facciamo accoppiare i figli saranno la 1° generazione ed è simboleggiata da F1, se fra questa generazione scegliamo una coppia e li facciamo accoppiare avremo la seconda generazione simboleggiata da F2 , poi la F3, la F4 e cosi via. Ma attenzione gli accoppiamenti fra fratelli non sono mai molto consigliati, no oltre massimo la F2 , perchè tendono a dare per via della consanguineità difetti nei figli, non solo morfologici ma anche biologici, cioè avremo Betta più deboli . Ogni selezionatore dovrebbe avere un quaderno, dove annota gli incroci che fa, cosi da non commettere errori, chiamando ogni esemplare con un codice dove ne è riportata la forma, la colorazione e la discendenza.
Prima di continuare voglio spiegare dei concetti che ci saranno utili:
Genotipo: è l’insieme di geni che compongono il DNA di un organismo, quindi il suo patrimonio genetico, comprendendo caratteri che manifesta e che non manifesta.
Fenotipo: è l'insieme di tutte le caratteristiche osservabili di un organismo vivente, cioè quello che noi vediamo nel suo aspetto fisico.
Omozigote: con gli alleli dello stesso gene (es. AA o aa)
Eterozigote: con gli alleli di diverso gene (es. Aa)
Iniziamo a parlare dei concetti base di genetica delle forme, fra i Show Betta che rappresentano il top della selezione abbiamo certamente gli HM e gli HM shortfin ( anche chiamati Plakat simmetrici) , voglio iniziare proprio citando questi perché in se rappresentano un carattere dominante e recessivo . Dovete per prima cosa tenere presente dei punti fondamentali con gli HM, per prima cosa che si sono ottenuti dei Betta con pinna aperta a 180°, e quindi raggi superiori dritti, superando la forza di gravità, nelle altre forme abbiamo aperture inferiori o con raggi curvati e cadenti, perchè “l’antigravità” (rappresentata dall’ HM) perde nei confronti della “gravità”, cioè delle code chiuse e cadenti.
Altro discorso recessivo, la simmetria è recessiva rispetto al “Caos” , la simmetria rappresenta un ordine di lunghezza fra le pinne, e questo lo si ha specialmente negli HM pinne lunghe e HM pinne corte, la cui caratteristica selettiva è appunto la simmetria fra le pinne, quindi tenete presente questo altro punto pratico.
Ultimo concetto base di selezione , e questo è un vero e proprio contrasto logico, le pinne lunghe sono dominanti sulle pinne corte , e anche questo lo spiegheremo in seguito, per ora tenete presente come concetti base queste 3 regole:
Pinne chiuse e cadenti vincono su pinne aperte (la gravità è dominante su l’antigravità)
L’asimmetria vince sulla simmetria (il disordine è dominante su l’ordine)
Le pinne lunghe dominano sulle pinne corte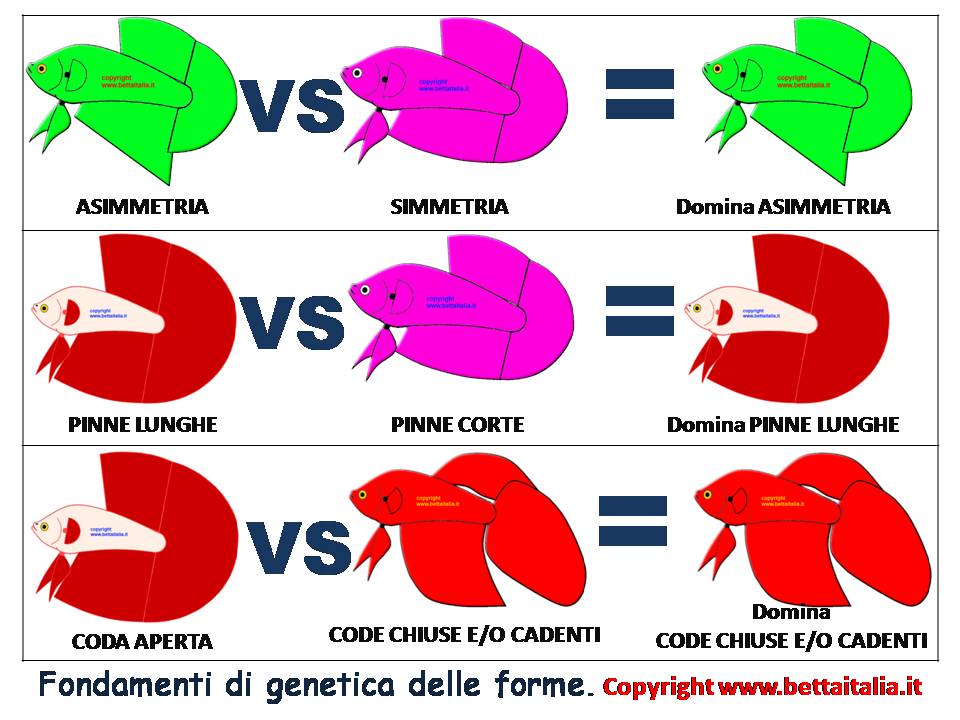
In questo schema vediamo i primi 3 esempi base: 1)Dominanza asimmetria, 2) Dominanza pinne lunghe , 3) Dominanza pinne cadenti.
Un’altro concetto che c’è da ricordare è che minori ramificazioni è dominante su maggiori ramificazioni, ma questo non vale sempre come vedremo, perchè per esempio il difetto Rosetail tende poi a tornare nelle generazioni.
Geno, cosa vuol dire? spesso leggete questa parola , HM geno o Dt geno ecc, bene Geno vuol dire che un betta ha determinati geni anche se fenotipicamente non li dimostra, per esempio HM geno DT sono degli HM che essendo figli di incroci fra HM e DT presentano caratteristiche HM ma con pinne più o meno voluminose , indice che hanno geni DT.

Betta splendens HM geno DT , si evidenzia che ha geni DT in particolare per la base della dorsale molto ampia
Fonte della foto: www.aquabid.com
Come vedete sto usando concetti semplici , perchè voglio andare incontro anche ai neofiti, in giro si trovano articoli sulla genetica con complessi schemi di Mendel , ma chi non ne è un po’ pratico poi poco ci capisce, mi ricordo la prima volta che vidi io articoli simili, alla fine ho capito più con anni ed anni di pratica.
Tornando al discorso, abbiamo citato i concetti base, ma vi direte, questi non comprendono i CT, i DT , o i nuovissimi Dumbo, ma io voglio arrivarci per gradi.
Iniziamo con esempi pratici, voglio iniziare proprio con i PK, perchè li ritengo più semplici, se accoppiamo un HMPK con un PK traditional cosa esce fuori? come abbiamo detto la gravità vince su l’antigravità e quindi essendo i Traditional PK avere una coda meno aperta e più arrotondata questo carattere vince sul carattere HM , quindi i Traditional PK li vedremo dominare sugli HMPK come forma della coda.
Altro esempio, un HMPK x HM shortfin , essendo entrambe con code HM questo carattere lo manterremo ma come abbiamo detto il disordine vince su l’ordine, quindi è dominante il carattere asimmetrico degli HMPK e tenderemo ad avere in maggioranza PK asimmetrici , potremo riselezionare dei PK simmetrici solo dalla F3 in poi .
HM x SD , i Super Delta sono dominanti sugli HM , bisogna vedere la discendenza dell’SD in questione, dato che spesso escono anche da accoppiamenti di Hm e vengono considerati scarti. Se è un SD geno HM avremo maggiori probabilità di ottenere figli HM , se invece è un’ SD puro o ancor peggio Deta geno o Vt geno , le probabilità diminuiscono. Normalmente da HM x SD si ha la quasi totalità di SD e qualche HM che in genere è di scarsa qualità, spesso si evidenzia la discendenza SD da una dorsale più piccola, meno aperta e cadente. Se vogliamo selezionare degli HM da questo incrocio possiamo tentare l’impresa, ma non ci darà mai HM di buona qualità, possiamo accoppiare fra loro i figli SD geno HM e dalla f3 in poi avere più HM che SD , ma poi a sfavore arriva il fattore consanguineità .
Abbiamo un discorso disastrante se invece facciamo gli incroci HM x VT o HM x Delta , la coda VT è la più cadente e questo è il fattore più dominante, perchè chiusa e cadente, cioè la massima espressione della forza gravitazionale, quindi da HM x VT avremo in maggioranza VT e solo qualche Delta o addirittura SD , ma con caratteristiche molto scadenti. Invece incrociando HM x Delta, come sappiamo Delta essendo una coda aperta non più di 120° ed essendo parente genetico stretto del VT, avremo in maggioranza dei Delta e qualche SD però come recessivi.
Riassumento in questo schema vediamo che : incrociando HM x SD avremo in prevalenza SD , invece incrociando HM x VT in maggioranza avremo dei Delta.
Accoppiando HM x HM cosa avremo? a questa domanda è difficile rispondere, perché bisogna vedere la discendenza degli HM, se hanno geni DT, o SD o OHM ecc, prendiamo il caso di HM veri e puri, quindi entrambe con coda perfetta a 180° , dovete sapere che anche accoppiando 2 caratteri antigravitazionali non daranno mai tutti figli cosi, infatti accoppiando 2 HM puri e senza geni DT o altro tendiamo ad avere una media di solo il 60-70% di HM , il resto saranno SD e in alcuni casi anche Delta, cioè ci sarà sempre un effetto specchio nelle generazioni che ci mostreranno da dove vengono gli HM , e cioè che sono frutto della selezione di SD ed altro, fino ad arrivare alla loro discendenza Delta. La percentuale di HM varia anche in base alla qualità selettiva e dunque alla purezza della forma, da una coppia di HM di bassa qualità tenderemo ad avere anche meno del 50% di HM , invece in una coppia di alta qualità potremo avere anche il 90% di HM, la purezza della forma è un traguardo per molti e anche mio, quindi la percentuale di HM media del 60-70% è solo indicativa.
Quindi dimenticatevi di avere tutti HM da un accoppiamento HM x HM . E qui voglio introdurre come fanno i selezionatori ad ottenere una maggioranza nettamente più elevata e quasi totale di HM , si incrociano OHMxHM .
Gli OHM sono Betta che presentano la coda con un’apertura maggiore ai 180° , quindi 200° e anche a volte oltre , questa super apertura da molti standard è considerato un difetto, ma dato che un’apertura maggiore tende ad essere recessiva e a perdersi nelle generazioni, accoppiando un HM che ha un’apertura maggiore con uno a 180° avremo circa il 70% di HM lineari e il resto OHM. Questo è un semplice modo per selezionare i migliori HM mantenendone le caratteristiche.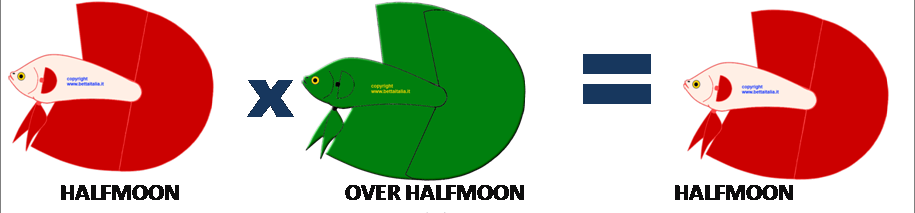
In questo schema vediamo un HM incrociato con un OHM (quello verde) , e da questo incrocio tenderemo ad avere maggiori HM.
Spesso invece specialmente in Europa si ha il brutto vizio di incrociare HM x DT , ma questo tipo di incrocio toglie le caratteristiche degli HM dandogli per esempio una dorsale maggiore, ma di questo ne parleremo più in la quando tratteremo la genetica dei DT. Si usano anche incroci fra HM e Rosetail per avere una percentuale più elevata di Hm , questi si tendono a dare più HM ma il difetto Rosetail poi tende a ripresentarsi nelle generazioni, quindi è sconsigliabile questo tipo di incrocio.
Ma geneticamente gli HM come possiamo definirli? io risalendo alla loro storia genetica li definisco semplicemente dei SD geno DT , perchè gli HM sono si frutto della selezione di SD accoppiando sempre gli esemplari che presentavano maggiore apertura, ma rimanevano sempre fortemente recessivi cosi la forma fu fissata incrociando questi primi HM che in realtà geneticamente erano sempre SD , con dei DT , quindi possiamo dire che geneticamente gli HM sono degli SD geno DT. Ed ecco perchè accoppiando 2 HM tendiamo a non avere tutta la prole HM ma anche degli SD, che come sappiamo è il gene dominante.
Continuiamo il nostro viaggio nella genetica pratica delle forme, abbiamo visto gli incroci fra pinne corte e fra HM, quindi riassumendo:
VT domina su tutti (VTxSD da in maggioranza VT e Delta, VT x Delta da in maggioranza VT)
Delta domina su SD e HM ma no su VT
RT (coda rotonda) domina su HM e SD ma è recessiva su VT e Delta
SD domina su HM ma no su Delta e VT
HM domina su OHM ma è recessivo su VT, SD e Delta
Ora vediamo gli incroci fra pinne lunghe e pinne corte, ricordo che le pinne lunghe sono dominanti sulle corte.
Perché spesso vengono fatti questo tipo di incroci, per esempio fra HM e HMPK? la risposta è perchè i PK tendono ad avere un corpo più massiccio e doppio, caratteristica che invece si tende a perdere negli HM , quindi per rinforzare la forma del corpo si fanno questi tipi di incroci.
Veniamo al primo caso, Traditional PK x HM , le pinne lunghe vincono, quindi è un carattere dominante che ha l’ HM ma il Traditional PK ha due fattori dominanti a suo favore, l’asimmetria e la coda arrotondata, che saranno trasmessi alla maggioranza della prole, quindi avremo dei Betta in prevalenza con le pinne lunghe o semilunghe , ma totalmente asimmetriche , tendenzialmente con anale più lunga e più triangolare che trapezoidale, con il lato posteriore più grande, e una coda che ricorda più un SD o in alcuni casi anche rotonda.
In questo schema si mette in evidenza la forma della coda in un'incrocio HM x TPK , avremo Betta con coda Delta o SD
I geni dominanti sono rappresentati in lettera maiuscola, invece quelli recessivi in minuscola. Il carattere dominante si manifesta fenotipicamente , nel caso di eterozigoti, es. Aa il gene recessivo (a) influenza il gene dominante (A) , cioè anche se si tenderà a manifestare fenotipicamente il carattere dominante, in parte della prole il carattere recessivo può manifestarsi più o meno parzialmente.
Ora per la prima volta faremo un'analisi genetica teorica utilizzando le famose tabelle di Mendel. Simboleggiamo ogni carattere con 2 lettere, se il carattere è dominante avrà lettere maiuscole, se è recessivo le avrà minuscole, se è eterozigone avrà una letterà maiuscola e una minuscola, se invece è omozigote avrà lettere uguali. Perchè 2 lettere per carattere? perchè gli alleli sono 2, procediamo con l'analisi in F1 :
Pinne lunghe : LL
Pinne corte: ll
Coda HM: hh
Coda inferiore ad HM : HH
Asimmetria: SS
Simmetria : ss
Genetica del Traditional PK: ll , HH , SS
Genetica HM: LL , hh , ss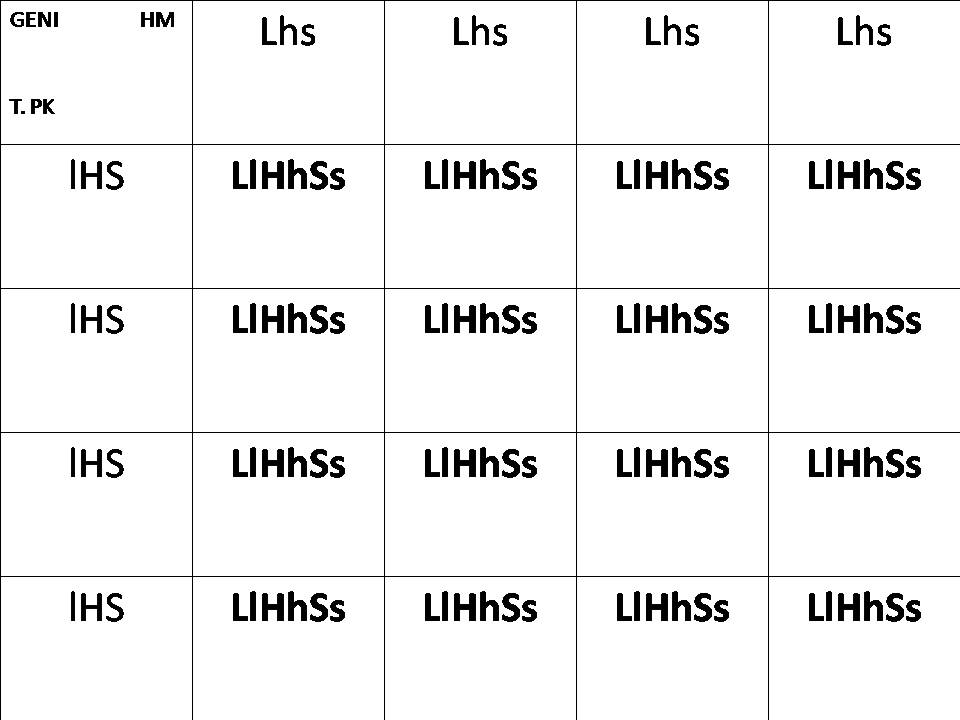
Risultato: LlHhSs (fenotipicamente si manifesteranno i caratteri dominanti , ma in parte della prole possono manifestarsi più o meno parzialmente i caratteri recessivi). Cioè avremo un Betta che fenotipicamente tenderà a presentare : Pinne lunghe, Coda SD o Delta, e pinne asimmetriche fra loro, in particolare con anale più lunga.
HM x HMPK , in questo incrocio tenderemo a non perdere la coda a 180° , avremo pinne lunghe in prevalenza, anche se io amo chiamarli mezze pinne, perchè ho più volte notato che la crescità generalmente non è proprio da veri HM pinne lunghe ma è un pò più contenuta, anche se fra i figli c’è chi raggiunge la lunghezza normale degli HM. Anche in questo caso 180°x180° ci daranno alcuni SD.
Facciamo l'analisi genetica teorica con le tabelle di Mendel in F1 anche in questo caso:
Pinne lunghe : LL
Pinne corte: ll
Asimmetria: SS
Simmetria : ss
Genetica del HMPK: ll , SS
Genetica HM: LL , ss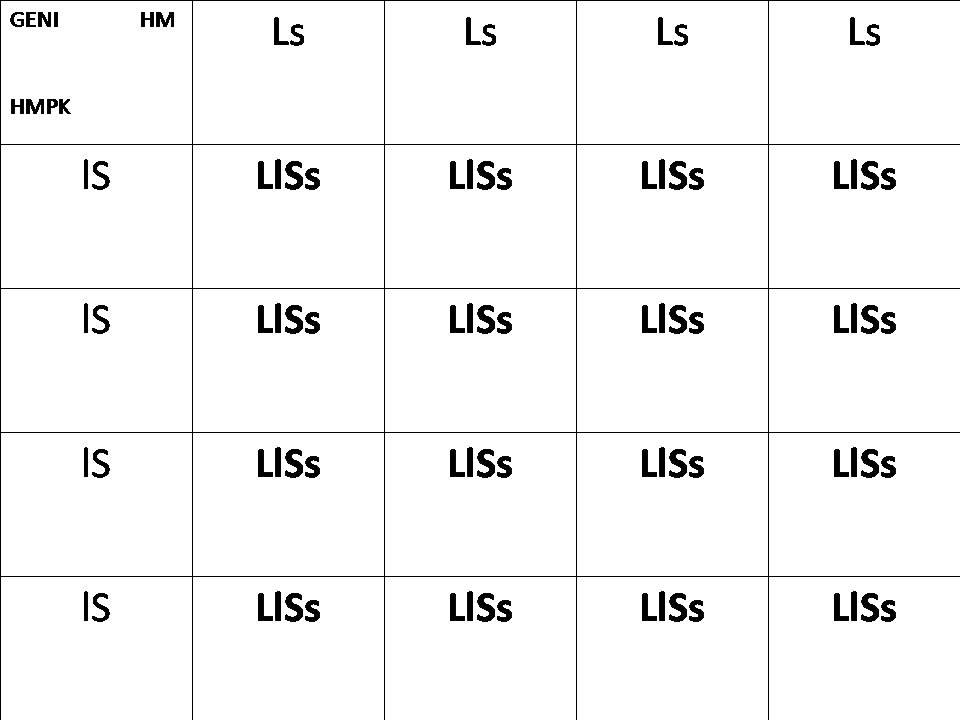
Risultato: LlSs, cioè Betta che tendenzialmente hanno le pinne lunghe o semilunghe , e in maggioranza asimmetriche con l'anale più lunga.
HM x HM shortfin , in questo caso abbiamo il carattere simmetrico in entrambe i casi , il carattere pinna lunga che domina e il carattere HM in entrambe i casi, quindi in prevalenza degli HM pinne lunghe tendenzialmente simmetriche , ma anche in questo caso avremo degli SD e dei Betta asimmetrici, perchè come abbiamo detto il carattere 180° e il carattere simmetria tende comunque a non essere totalitario e a perdersi essendo recessivo.
Io non amo questo genere di incroci perchè ho più volte notato che il carattere pinne corte crea grande disturbo poi nelle discendenze, avendo in prevalenza Betta HM che non sviluppano al 100% la lunghezza delle pinne come dovrebbero ma tendono a rimanere limitate.
Ora passiamo ad esaminare i famosi Crown , qui ci addentriamo in un discorso alquanto complesso.
Quando si parla di CT bisogna capire a quale si riferisce, i migliori CT come sappiamo sono quelli che presentano un’apertura della coda a 180° , quindi come sappiamo è recessivo e spesso per selezionare CT cosi si incrociano con gli HM, facendo delle selezioni specifiche.
Per prima cosa teniamo presente che il CT è recessivo ma no totale, lo considero un recessivo parziale , inquanto i suoi geni lasciano sempre la loro presenza ben visibile.
Poniamo i primi casi, accoppiamo un HM con un CT 180° cosa esce?
Per la prima volta facciamo un’analisi carattere per carattere:
Chiamiamo il carattere pinne integre con II e non integre con ii , dall’incrocio di questi geni abbiamo tutti figli Ii cioè dei Geno CT , chiamati anche Combtail o Halfsun. Quindi da un incrocio fra HM x CT avremo degli Halfsun e dei Combtail, cioè gli HS sono quelli che presenteranno apertura a 180° ma con una semi riduzione della membrana non superiore al 30% , cioè è un segno che ha geni CT , quindi come abbiamo detto si chiamano anche HM geno CT . Ma come sappiamo il gene 180° non si trasmette a tutti i figli, quindi una parte saranno degli SD geno CT e forse anche qualche Delta geno DT, cioè con una coda che ha apertura inferiore a 180° ma che presentano una lieve riduzione della membrana con i raggi che escono (Combtail). Ma attenzione in questi incroci non è detto che tutti i geno CT che nasceranno mostrino tutti lo stesso livello di raggi fuoriuscenti dalla membrana, infatti anche essendo tutti geno CT , ognuno manifesterà questa caratteristica in modo diverso, per esempio alcuni possono anche non manifestarla, o manifestarla molto lievemente cosi da sembrare al 100% degli HM, o altri manifestare anche una riduzione della membrana cosi eccessiva da sembrare al 100% CT.
1) 2)
2)
Foto 1: Combtail. Foto 2: HalfSun. Fonte della foto 1 google immagini , fonte della foto 2 www.aquabid.com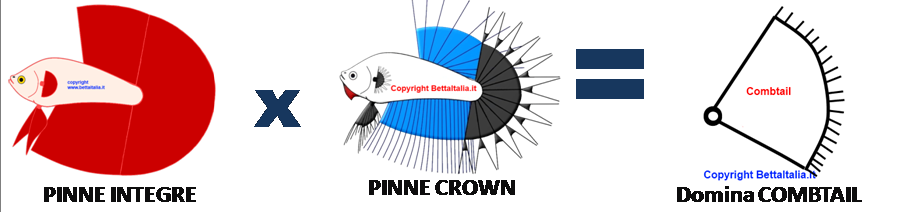
Rappresentazione di un' incrocio fra un HM e un CT , risultato in maggioranza Combtail
Questo tipo di incroci io li chiamo “contenitori genetici” , perchè all’interno dei figli ci sono entrambe i geni HM e CT, e questi si manifesteranno delle discendenze , se faremo accoppiare un HM geno CT con un’ HM avremo maggiori figli HM , perchè verrà trasmesso il carattere HM in prevalenza, e se invece lo accoppieremo con un CT potremo selezionare o perfezionare una linea CT a 180° , quindi come incroci di transizione per migliorare specialmente linee CT sono molto utili.
Analisi genetica teorica con tabelle di Mendel in F1, incrociando un CT ipoteticamente con apertura SD con un' HM
Pinne integre : II
Pinne crown: ii
Genetica dell’ HM: II
Genetica CT: ii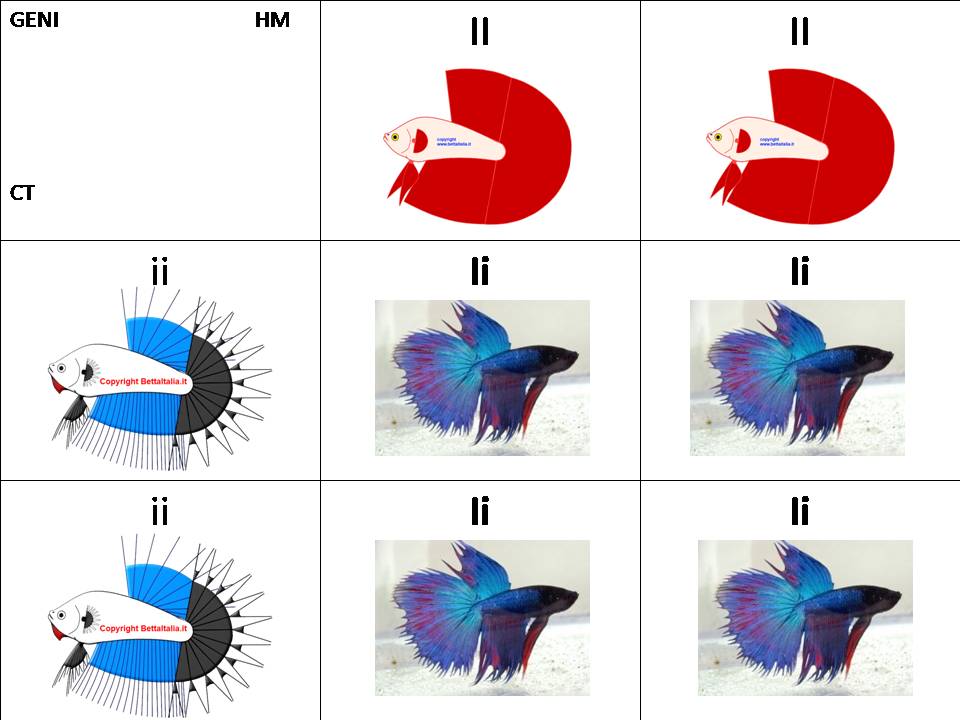
Risultato: Ii , cioè un Betta geno CT , che può essere HS o Combtail
Incrociando la F1 con i geni Ii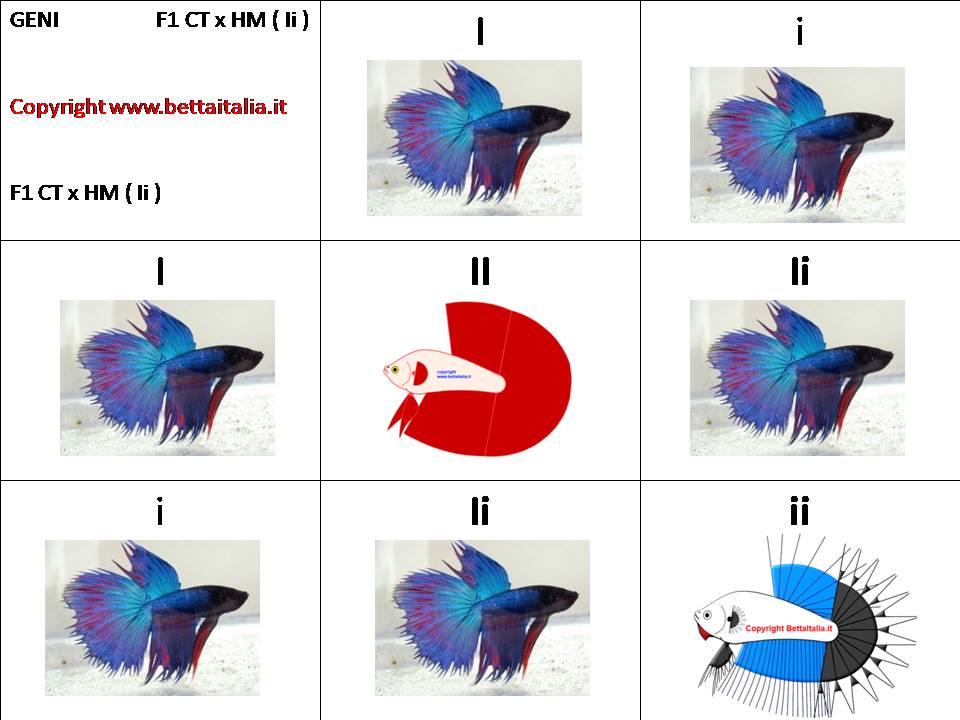
In F2 avremo: il 25% di figli II che presenteranno pinne completamente integre, il 50% di Combtail (Ii) e il 25% di ii cioè di CT
Analizziamo altri tipi di incroci:
CT x VT , qui per prima cosa vincono le pinne cadenti del VT , i figli saranno dei VT geno CT comunemente chiamati Pinne a Pettine.
CT x SD , dipende da che CT è, ma presumiamo che sia un 180° , come sappiamo vince il gene SD , quindi non avremo figli con apertura a 180° , ma avremo degli SD geno CT chiamati anche Combtail. Se il CT invece ha apertura a Delta avremo dei Delta geno CT
Delta x CT, se il CT ha pinne a velo come apertura usciranno dei VT geno CT , se il CT ha apertura 180° , SD o Delta avremo in maggioranza dei Delta geno CT chiamati anche Combtail
Ricordo che facendo incroci per formare e purificare una forma ci vogliono almeno 3 generazioni.
Ora vediamo gli incroci fra CT e Betta a pinne corte:
CT x Traditional Pk , in questo caso avremo dei Combtail, essendo il carattere pinne corte recessivo e CT recessivo, i figli in prevalenza prenderanno il gene pinne lunghe, come apertura della coda se il CT ha l’apertura della coda a VT naturalmente vincerà su l’apertura dei traditional PK , quindi avremo dei pesci con pinne lunghe o semilunghe , con presente più o meno riduzione della membrana nelle pinne e apertura tendenzialmente al Delta.
CT x HMPK , spesso discorso fatto sopra, ma qui abbiamo il carattere HM nel PK , quindi se anche il CT è 180° avremo degli HalfSun con pinne più o meno lunghe.
CT x HM shortfin , qui abbiamo un discorso differente , abbiamo simmetria per simmetria, quindi se invece nei casi di sopra possiamo avere tendenzialmente asimmetria nelle pinne qui invece conserviamo una certa simmetria , quindi senza ripetere quello detto sopra su apertura della coda e lunghezza delle pinne, avremo dei Halfsun simmetrici se il CT è 180°
Esempio: l’incrocio di PK in generale con dei CT (siano essi sd, vt ecc, non analizzeremo l’apertura delle code come sopra ma solo come si comporta il gene CT). Quindi accoppiamo un qualsiasi pinna corta con un CT pinne lunghe , PK x CT , avremo dei figli geno CT , cioè Combtail pinne lunghe . E se vogliamo selezionare dei PKCT? ricordate che per creare e fissare una forma ci vogliono sempre almeno 3 generazioni , dalla F1 avremo appunto questi geno CT , accoppiando 2 di questi figli in F2 avremo circa il 6,25% di Betta PKCT, cioè che presenteranno corte e CT, bene per fissare questa forma li dovremo accoppiare e in F3 avremo il 100% di PKCT. Ma questa ricordatevi che è teoria, ogni esemplare ha una sua storia genetica. Infatti per esperienza c’è quando le cose sono state più facili perchè erano esemplari geneticamente più predisposti, e quando è stato un lavoro più difficile.
Rappresentazione genetica con tabelle di Mendel di un'incrocio fra PK e CT in F1
Pinne lunghe : AA
Pinne corte: aa
Pinne Integre: BB
Pinne Crown : bb
Genetica del PK: aaBB
Genetica CT: AAbb 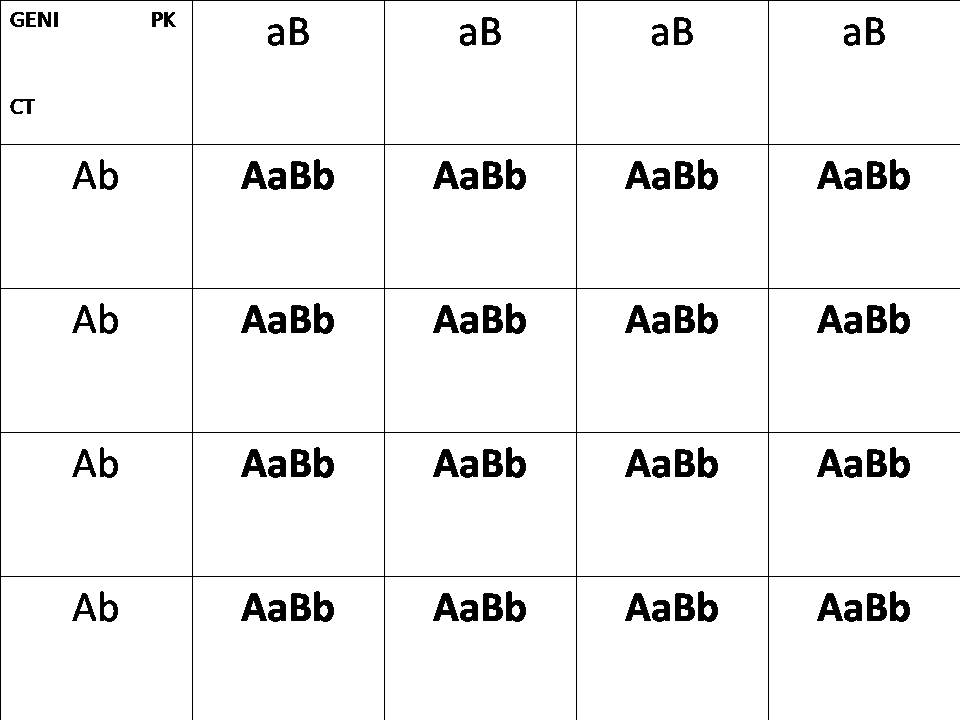
Risultato in F1: AaBb , risultato un combtail pinne lunghe
Incrociamo la F1 AaBb X AaBb
Avremo F2: 
AABB e AaBB che avranno normali pinne lunghe integre
AABb e AaBb che saranno normali Combtail pinne lunghe
AAbb e Aabb che saranno Crown pinne lunghe
aaBB che saranno normali PK
aaBb che saranno dei PK combtail. 
aabb che saranno i veri CTPK.
In F3 ci basterà accoppiare aabb x aabb ed avremo tutti PKCT
Ora prima di passare ad analizzare il singolare caso dei DT voglio aprire una piccola parentesi che riguarda i famosi Big Ear e Dumbo , una delle ultime selezioni.
Il Big Ear è caratterizzato come sappiamo da grandi pinne pettorali, iperamificate , sono una forma rosetail delle pettorali, le ultime selezioni chiamate Dumbo le hanno ancora più grandi. Questo carattere è recessivo, quindi se accoppiamo un Betta con pettorali normali con un Big Ear la maggior parte dei figli non mostrerà pettorali grandi , ma saranno portatori del gene , e potremo riottenere questa caratteristica in f2 o f3. Tendenzialmente si trovano Big Ear PK e no HM , perchè i Big Ear sono anche portatori del gene Rosetail , il quale tende a non manifestarsi in Betta a pinne corte , ma se selezioniamo degli HM Big Ear questi tenderanno a soffrire di pinne a Rosetail anche eccessive, e in più avremo un Betta con le pinne molto pesanti. I difetti morfologici di cui sono portatori sono: corpo più piccolo, dorsale più piccolo e pinne rosetail accentuate, difetti che tendono a non manifestarsi nei PK Dumbo , perchè i Pk sono portatori di un corpo più massiccio , e tendono a non manifestare il rosetail nelle pinne avendole corte.
Analisi genetica con le tabelle di Mendel fra un non Dumbo e un Dumbo
Pettorali normali : AA
Pettorali Dumbo: aa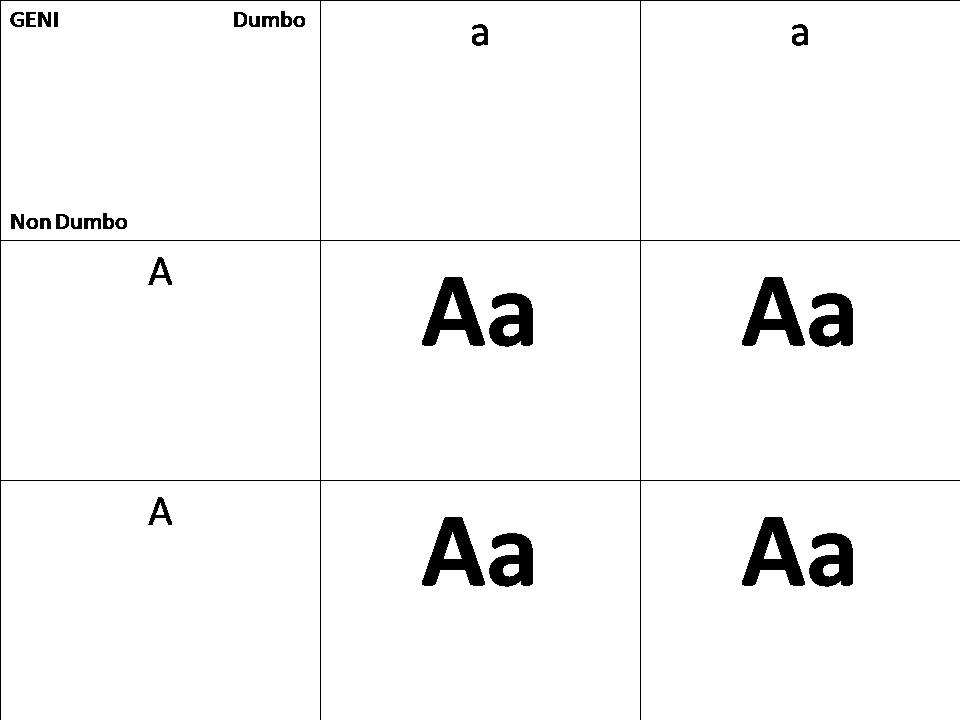
Risultato in F1: Aa , risultato un Betta non Dumbo ma che potrebbe manifestare delle pettorali leggermente più grandi in alcuni individui. Chiamati Betta geno Dumbo
Accoppiamo 2 figli della F1 Aa X Aa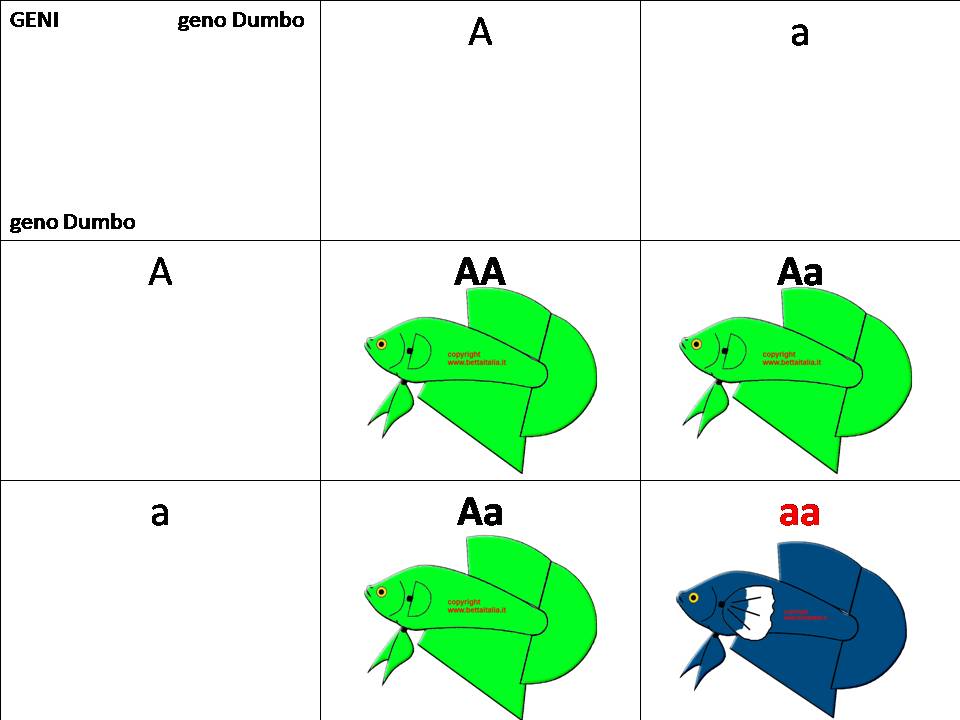
Avremo in F2:
AA e Aa che saranno normali Pk non Dumbo
aa che invece saranno Dumbo
In F3 ci basterà accoppiare aa x aa ed avremo tutti Dumbo
Altra piccola parentesi per i famosi Giant, che appena usciti ebbero tanto successo oggi invece meno diffusi e ricercati. Sappiate che anche il gene Giant è recessivo. Possiamo paragonarlo al gene Dumbo, perchè si comporta circa allo stesso modo.
E ora finalmente trattiamo quel vero rompicapo che è il gene DT , e cioè i Betta che presentano 2 lobi caudali, uno sopra e l’altro sotto e anche una dorsale più grande. Leggevo recentemente su un forum asiatico che stando ad alcuni recenti studi genetici sui Betta DT questi in realtà non sarebbero altro che frutto di una malformazione gemellare, cioè i primi Betta DT da cui poi sono stati selezionati tutti i DT, non era altro che un uovo che al suo interno aveva 2 gemelli, ma nello sviluppo i 2 embrioni si erano fusi, come accade spesso nelle malformazioni gemellari, fusi in un solo corpo ma avendo il pinnaggio equivalente a 2 Betta , ed ecco perchè un corpo più grosso e una dorsale e anale grandi il doppio oltre che a 2 lobi caudali, sarà vero? può esserlo, ma non ho mai trovato fonti per accertare questa cosa.
Per prima cosa il gene DT è recessivo, ma è proprio grazie a un DT che oggi abbiamo gli HM , si perchè i DT trasmettono l’ampiezza delle pinne anche se invece il carattere doppia coda è recessivo.
Di DT come sappiamo ne abbiamo vari tipi, prendiamone sono in esempio 2 tipi i VTDT chiamati anche coda di rondine e gli HMDT , cioè Betta con 2 code simmetriche la cui unione danno i 180°
Abbiamo anche i PKDT gli SDDT ecc, ma qui ora vediamo solo come si trasmette il gene doppia coda e cosa comporta, il discorso legato a pinne lunghe e corte , o a poca e polta apertura della coda, qui non li trattiamo dato che sono stati trattati prima, poi vedremo anche come si comportano i 2 geni recessivi CT e DT se vengono incrociati.
Per prima cosa voglio sfatare un mito, almeno nel mio piccolo, più volte leggo e mi è stato detto che 2 DT non si possono far accoppiare perchè i figli sarebbero malformati....., a quanti accoppiamenti ho fatto cosi non mi è mai capitato un figlio deforme, sarò stato fortunato io? ma anzi con queste selezioni ho migliorato la simmetria fra le 2 code. Non voglio dire che questa affermazione sia sbagliata, ma spesso si prendono per vere le cose scritte da altri, e secondo me andrebbe tutto testato, a me non sono mai capitati deformi, ne mai sul web ho visto figli di 2 DT nati deformi, a me mai successo nulla di negativo. Ma c’è da dire che sicuramente questo è dovuto dal fatto che difficilmente si trovano dei DT puri omozigoti (cioè con entrambe gli alleli DT ) , ma quasi tutti sono quelli che io chiamo falsi DT , cioè geneticamente eterozigoti con geni HM e DT ma che manifestano le 2 code. Unico difetto riscontrato incrociando 2 DT è stato pinne un pò più ramificate in alcuni figli, e naturalmente più pesanti perchè voluminose, ma solo in alcuni figli.
Veniamo a degli esempi :
HMxDT , geneticamente avremo tutti degli HM geno DT , perchè il gene a singola coda domina, per quanto riguarda l’apertura dipende il DT che code ha , se sono VT, SD ecc, questo discorso lo abbiamo già fatto , come sappiamo vince la pinna lunga e meno chiusa. Anche se per ipotesi dovremmo avere Betta più simili a un’ HM che a un DT , in realtà avremo dei Betta in F1 HM ma che presentano le pinne più ampie e in particolare la dorsale, infatti è un carattere dei DT che tende a rimanere , poi avremo dei Betta che mostreranno una sorta di doppia coda, cioè una divisione non proprio netta o ancora non simmetrica, cioè vedremo peresempio delle code con 2 lobi ma con uno dei 2 molto più piccolo, e solo qualcuno potrà presentarsi uguale o simile a un’ DT. Molti si staranno dicendo: questa cosa non è vera perchè io ho accoppiato HM e DT e mi sono usciti metà DT veri, infatti come ho detto il risultato è solo teorico, perchè dipende dalla genetica dei nostri HM e dei nostri DT, purtroppo spesso gli HM hanno geni DT perchè figli o nipoti di accoppiamenti con DT , quindi spesso in accoppiamenti fra HM e DT non ho riscontrato i risultati teorici secondo la tabella di Mendel, anche per questo sono sfavorevole agli incroci con DT a fini selettivi della linea HM. Come ho detto sopra , gli HM conservano geni DT perchè la forma fu selezionata grazie ad incroci con essi , per questo io li definisco geneticamente degli SD geno DT , ed ecco che spesso negli incroci abbiamo anche vari DT veri e propri.
Esempio di Hm geno DT che manifesta 2 code asimmetriche, quella di sopra in questo caso è molto più piccola.
Facciamo l'analisi genetica con le tabelle di Mendel fra un Doppia Coda e un Coda Singola
Coda Singola : AA
Doppia Coda: aa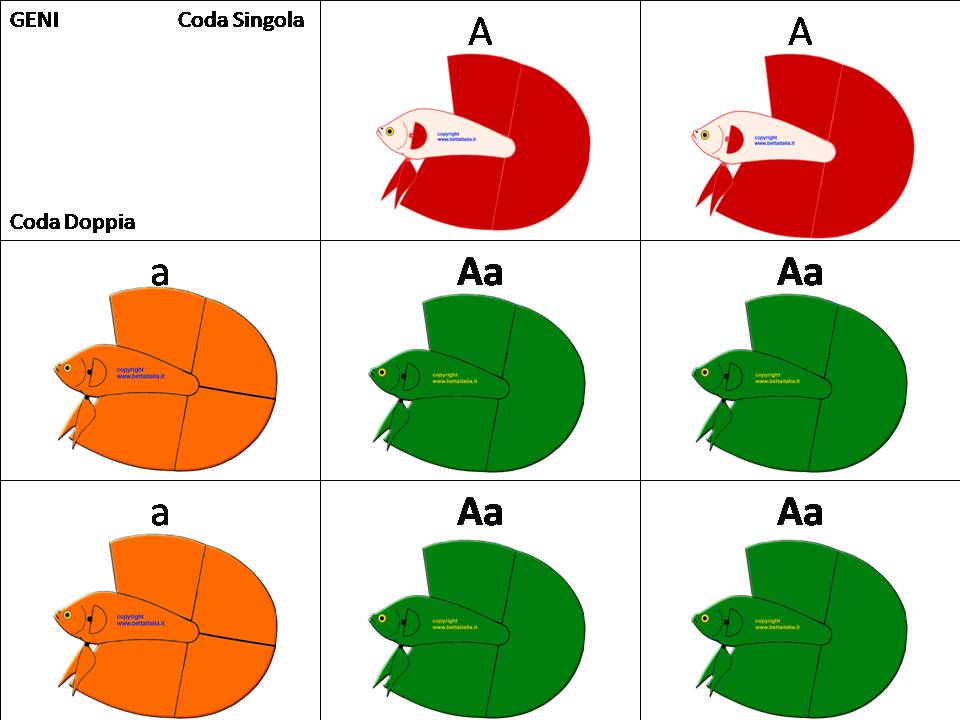
Risultato in F1: Aa , risultato un Betta Hm geno DT , che cioè teoricamente dovrebbe presentare una sola coda e pinne più voluminose, ma come abbiamo detto nascono anche DT asimmetrici e forse qualcuno simmetrico.
Ora incrociamo la F1 Aa X Aa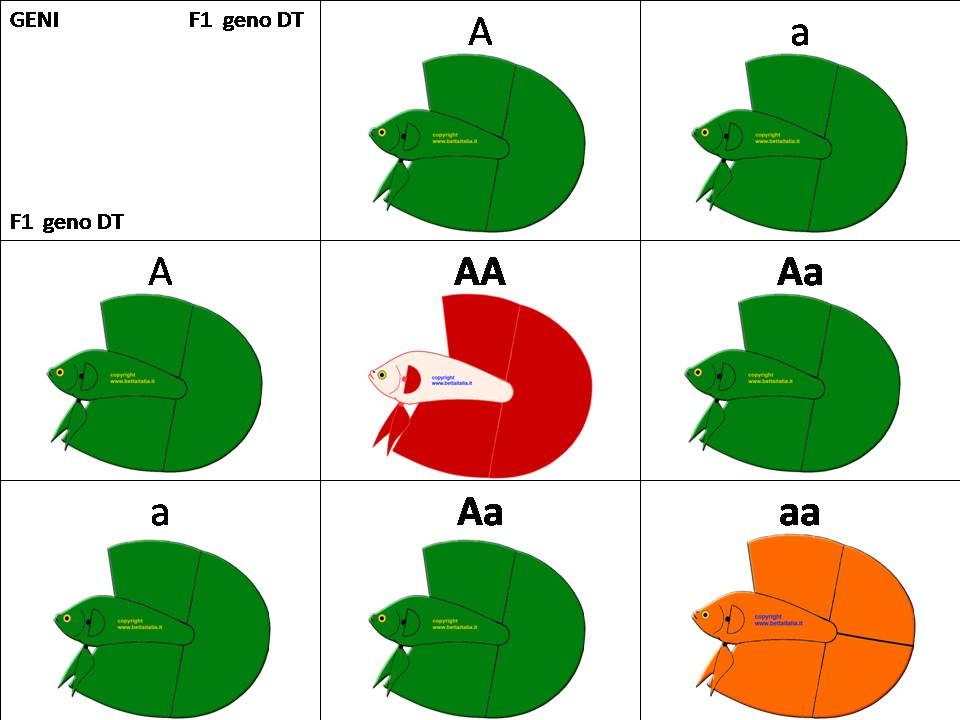
Avremo F2: AA che saranno normali Coda Singola. Aa che sono i geno DT . aa che saranno dei DT puri
In F3 ci basterà accoppiare aa x aa ed avremo tutti DT (anche se come abbiamo detto alcuni allevatori questo incrocio non lo consigliano)
Ora prendiamo per esempio l’incrocio di PK in generale con dei DT (siano essi sd, vt ecc, non analizzeremo l’apertura delle code come sopra, ma solo come si comporta il gene DT). Quindi accoppiamo un qualsiasi pinna corta con un DT pinne lunghe , PKxDT , avremo dei figli PK geno DT , cioè con delle pinne lunghe e semi lunghe e più ampie e ramificate, in primis la dorsale . E se vogliamo selezionare dei PKDT? ricordate che per creare e fissare una forma ci vogliono sempre almeno 3 generazioni , dalla F1 avremo appunto questi geno DT , accoppiando 2 di questi figli in F2 avremo circa il 6,25% di Betta PKDT, cioè che presenteranno pinne corte e 2 code, bene per fissare questa forma li dovremo accoppiare e in F3 avremo il 100% di PKDT . Ma questa ricordatevi che è teoria, ogni esemplare ha una sua storia genetica. Infatti per esperienza c’è quando le cose sono state più facili, perchè erano esemplari geneticamente più predisposti e quando è stato un lavoro più difficile.
Facciamo l'analisi genetica con le tabelle di Mendel fra un PK Doppia Coda e un PK Coda Singola:
Pinne lunghe : AA
Pinne corte: aa
Coda singola: BB
Coda doppia : bb
Genetica del PK: aaBB
Genetica DT: AAbb 
Risultato in F1: AaBb , risultato un geno DT pinne lunghe e semilunghe. Cioè che presentano in particolare una dorsale più voluminosa e una coda singola o 2 fortemente asimmetriche.
Ora incrociamo la F1 AaBb X AaBb
Avremo in F2 il 6,25% di aabb che saranno veri PKDT
PKDT
In F3 ci basterà accoppiare aabb x aabb ed avremo tutti PKDT
Una piccola parentesi, accoppiando VTxDT io ho ottenuto varie volte dei Super Delta.
Ora l’ultimo esempio prima di concludere, incrocio fra CT e DT. Non prenderemo in caso l’apertura delle code , ma prendiamo come obbiettivo quello di voler fare dei doppia coda CTDT , come dovremmo in via teorica fare?
Sappiamo che l’effetto Crown è recessivo ma anche il Doppia Coda è recessivo , bene accoppiamo dei CT x DT , prima generazione avremo un qualcosa di veramente strano, che dipenderà molto dall’apertura delle code dei genitori, quindi diciamo che avremo in linea generica dei Combtail a pinne ampie, cioè dorsale più ampia e coda maggiore.
Quindi avremo questi Betta a coda singola ma con questi raggi semiusciti nelle pinne, bene continuiamo.
Accoppiamoli in F2 e avremo circa il 6% di veri DTCT , accoppiamo questi esemplari fra loro e in F3 avremo fissato la forma.
Facciamo l'analisi genetica con le tabelle di Mendel fra un CT e un DT:
Pinne integre : AA
Pinne Crown: aa
Coda singola: BB
Coda doppia : bb
Genetica del CT: aaBB
Genetica DT: AAbb 
Risultato in F1: AaBb , risultato un geno DT combtail , con le pinne più voluminose.
Ora incrociamo la F1 AaBb X AaBb
Avremo F2 dei Betta che presenteranno solo code singole, altri solo code doppia, altri che saranno o geno CT , o geno DT o entrambe. Solo una minoranza sarà aabb e cioè veri CTDT Betta splendens CTDT. Fonte della foto: google immagini
Betta splendens CTDT. Fonte della foto: google immagini
In F3 ci basterà accoppiare aabb x aabb ed avremo tutti CTDT
Fra tutte le forme viste possiamo dire che la dominante in assoluto è la forma VT , cioè pinne lunghe, poco aperte e cadenti , questa forma vince su tutte anche su quelle a pinne corte. In natura i Betta splendens sono a pinne corte, e le prime selezioni ai primi del 900’ diedero i pinne a velo, infatti la condizione in natura ad avere le pinne corte è una condizione forzata , se tenuti in cattività in poche discendenze tendono subito a manifestare pinne più lunghe, ma in natura un Betta a pinne lunghe è svantaggiato, primo nel nuoto e secondo è più visibile ai predatori, quindi la selezione naturale costringe il Betta splendens in natura a rimanere con le pinne corte. Quindi i VT sono dominanti perchè la possiamo considerare la vera forma dei Betta splendens, che riescono però ad esprimere solo se allevati in cattività.
Ora alcuni cenni su come selezionare alcune forme più “pregiate” e qui riprenderò alcuni concetti detti sopra.
Selezioniamo gli HM
Come abbiamo detto accoppiando 2 semplici HM non tutti i figli saranno HM , e la percentuale si abbasserà se continuiamo ad accoppiare fra loro quelli che nasceranno HM , per prima cosa tenete in mente che gli accoppiamenti fra fratelli vanno fatti molto poco perchè sono controproducenti, a parte che i figli nascono più deboli, ma se accoppiamo oltre la F2 fra fratelli tenderemo a rovinare le linee, io generalmente accoppio fra fratelli solo la f1 , ma al massimo massimo consiglio di non andare oltre la F3 . Se accoppiamo spesso fra fratelli i caratteri recessivi come la coda più chiusa e la corsale più piccola saranno sempre più presenti nelle generazioni.
Questo lo potremo notare su quello che è il vero vantaggio degli HM, e cioè avere il primo raggio superiore della coda completamente alzato e dritto, potremo vedere come facilmente i figli dei figli presenteranno questo raggio curvato o non completamente a 180° , perchè appunto deve vincere la forza di gravità.
Quindi come possiamo selezionare ottimi HM? ci sono 3 soluzioni: accoppiarli con i DT o con i Rosetail o con gli OverHM.
Veniamo al primo caso, l’incrocio fra HM e DT , molto in voga in Europa , è giusto farlo? assolutamente no! molti lo fanno anche per dare una dorsale più ampia e un’apertura sicura a 180° della coda, ma chi fa questi incroci non imparerà mai nulla perchè è come prendere una scorciatoia, ed ottenere degli ibridi (di forma) che non hanno nessuna forma a se. Io sono un purista della forma, e vedere anche questi ibridi partecipare ai campionati e magari vincere ritengo che non si premia il vero lavoro di selezione, perchè sono i classici incroci da neofita che non sa selezionare. Quindi non fatelo! questo tipi di incroci danno anche nelle generazioni problemi ossei se fatti spesso, oltre che a code deformate nella forma, come orli ondulati, o con raggi più lunghi e altri più corti, e no il perfetto mezzo cerchio come richiesto. Personalmente amo di più le altre 2 possibilità, infatti possiamo incrociare un HM con un Rosetail , come sappiamo la poca ramificazione domina sulla molta ramificazione , quindi fra uno con ramificazione normale HM con uno avente una ramificazione eccessiva HM , avremo dei Betta HM più o meno con una buona ramificazione, mantenendo cosi sempre i 180° della coda. Ma questo tipo di incrocio ha i suoi lati negativi, spesso i Rosetail (non è sempre cosi dipende dalla genetica che ha) tendono poi a dare nelle discendenze dei corpi piccoli o anche delle dorsali minute, dipende da Rosetail a Rosetail, non è sempre cosi, ma questi sono i punti a sfavore di questo tipo di incrocio, e qui se si è dei bravi selezionatori ci si deve mettere a lavoro. Per prima cosa possiamo riavere dei corpi più grandi incrociando questi a loro volta con degli HMPK o ancora meglio degli HM shortfin , e se la dorsale è troppo piccola in questo caso ci può essere utile un incrocio con un DT , ma che non deve essere ripetuto per almeno 3 generazioni, io consiglio almeno 5.
Ecco l’ultimo possibile incrocio che possiamo fare, e che io amo di più , lo adottano anche la maggior parte dei selezionatori asiatici , è la selezione che ha meno lati negativi, anzi direi quasi nessuno se non si vuol essere pignoli, mi riferisco all’ incrocio fra HM e OHM . Se gli OHM non hanno geni Rosetail come spesso accade, ma hanno una bella coda con la membrana ben tirata e che si apre oltre i 180° e come loro caratteristica hanno anche una dorsale con un’apertura più ampia anche in avanti, con il primo raggio obliquo in avanti, questo tipo di OHM ha dei geni DT che non si manifestano in quanto pochi e non di fresco incrocio, cosi abbiamo una coda più aperta . Una coda con più apertura e quindi più raggi e membrana X una coda con la giusta membrana e raggi darà degli HM perfetti e qualche OHM che potremo utilizzare in futuri incroci, questo tipo di selezione della forma HM secondo me è la migliore ed è quella che suggerisco. Infatti quando compro per importazione dei show Betta cerco sempre di prendere una coppia con un HM e l’altro OHM. Generalmente le femmine HM pure non manifestano la coda a 180° ma la manifestano le femmine OHM. Un’altra possibile selezione di discreti HM è incrociare buoni SD , magari figli da accoppiamenti HM x HM , con un OHM , quindi quelli che nasceranno saranno degli SD geno OHM geneticamente ma fenotipicamente in maggioranza HM , unica pecca è che tendenzialmente avremo pesci con una dorsale con poca apertura, e spesso anche un’anale troppo lunga. Rimane il fatto che la crescita dei raggi, e dunque di tutte le pinne specialmente negli HM, è molto influenzata dall’acqua e quindi dall’ambiente in cui crescono , e questo lo vediamo fra quelli nati in acque Europee, che sono spessissimo soggetti a difetti. Quindi come ho già detto, la genetica ha il suo ruolo ma anche come e dove i Betta vengono allevati.
Selezioniamo i CT
Nel selezionare i CT dobbiamo considerare principalmente 2 fattori apertura della coda a 180° e raggi duri e dritti, la migliore selezione è accoppiare fra di loro solo gli esemplari con le migliori caratteristiche, ma vediamo come possiamo fare delle selezioni per migliorare per esempio un classico CT commerciale. Ipotizziamo che al negozio compriamo un CT commerciale che non sia proprio scadente e quindi con la coda cadente, ma con una coda ad apertura SD e i raggi fini. Prima selezione che va fatta è incrociare il nostro CT con un HM a pinne corte , che sia esso PK o HM shortfin , meglio il secondo. Perchè l’incrocio con un HM a pinne corte? per prima cosa sappiamo che il gene pinne lunghe è dominante , seconda cosa dobbiamo darli più apertura e terza cosa per avere dei raggi più grossi e doppi, e il miglior incrocio in questo caso è con i PK . In F2 otterremo degli esemplari con un’apertura molto simile ai 180° e con raggi duri ma che escono poco dalla membrana, e da questi potremo partire. E se accoppiamo il nostro CT con un HM pinne lunghe? certamente nella selezione ci guadagniamo in lunghezza delle pinne e apertura della coda, ma avremo figli con raggi troppo fini . Ricordatevi che la doppiezza e rigidità dei raggi nei CT ce le da l’incrocio con i PK. Senza addentrarmi molto nella genetica delle forme CT , spesso mi viene chiesto come si fanno ad ottenere le forme a raggi incrociati , come i King. Per prima cosa bisogna sfatare un mito, quel tipo di forma è una mutazione genetica e non è indotta dall’allevamento, o almeno non in tutti i casi. Possiamo facilmente ottenere i primi Betta a raggi incrociati selezionando incroci VT x CT 180° , il gene VT da raggi ricurvi e cadenti che andranno ad incrociarsi con quelli che rimarranno dritti, non entro molto nel discorso , ma questo giusto solo per spiegare che sono frutto di selezioni. Personalmente sto selezionando una forma di raggi incrociati che ormai si è persa, e sono quelli con i raggi ricurvi internamente.
Due parole sui PK
Penso che sulla selezione dei PK ci sia poco da dire , primo perchè sopra abbiamo citato i punti più fondamentali e secondo perchè la loro selezione è estremamente semplice, tanto che li consiglio ai neofiti per iniziare . Non hanno le difficoltà selettive dei CT o degli HM , che devono rispettare una certa lunghezza di pinne, una certa simmetria e i vari problemi strutturali delle pinne. E’ nettamente più facile selezionare un buon PK che un buon HM o CT . Infatti spesso chi da neofita parte subito con HM poi finisce che fa solo disastri e si annoia , bisogna prima fare pratica con i PK che sono veramente Betta da neofiti. E poi questi pesci sono proprio più forti e resistenti.
Creiamo i Dumbo HM
Ora proprio per concludere, prendiamo in ipotesi un ultimo esempio, facciamo caso che vogliamo creare un Dumbo HM , quindi un HM a pinne lunghe ma che presenti le pettorali più grandi, sia se siano Big Ear che sono la prima selezione e presentano pettorali grandi il doppio del normale, sia se siano Dumbo che sono la selezione più recente e presentano pettorali grandi più del doppio.
Come sappiamo in commercio sono più diffusi i Dumbo PK , perchè meglio si adattano a quella forma, purtroppo il gene Dumbo come difetto negli HM da pinne Rosetail, perchè le pettorali grandi non sono altro che pettorali Rosetail, essendo estremamente ramificate. Per questo gli HM Dumbo non sono molto amati e non è ritenuta una buona selezione.
Incrociamo il nostro PKDumbo xHM , i figli saranno HM genoPK e geno Dumbo, cioè in prevalenza saranno normali Hm qualcuno potrà mostrare pinne un pò più piccole e qualcuno pettorali un pò più grandi, anche se è raro. Incrociamo questi figli e in F2 con un pò di fortuna otterremo i primi HM Dumbo.
Analisi Genetica:
Pinne Lunghe : AA
Pinne Corte: aa
Pettorali normali: BB
Pettorali Dumbo : bb
Genetica HM: AABB
Genetica PK Dumbo: aabb
Risultato in F1: AaBb , risultato HM pinne lunghe o semi lunghe , tendenzialmente con pettorali normali, ma qualcuno può presentare pettorali leggermente più grandi.
Incrociamo la F1 AaBb X AaBb
Avremo F2 dei Betta che presenteranno solo code corte, altri solo code doppia, dei PK Dumbo e altri solo geno Dumbo. Solo una minoranza sarà AAbb e cioè veri HM Dumbo
Hm Dumbo.
In F3 ci basterà accoppiare AAbb x AAbb ed avremo tutti HM Dumbo. Anche i portatori dei geni Aabb si manifesteranno come HM Dumbo
Conclusioni.
Come sempre quando scrivo un articolo ci metto varie settimane, perchè amo scriverlo basandomi solo sulle mie esperienze e conoscenze, in giro fra i siti di acquariofilia e non solo di bettofilia , si trovano vari articoli sulla genetica delle forme, ma chi gli articoli li conosce si accorge che sono copiati o in parte copiati, e tradotti da famosi articoli in Inglese, e che spesso non sono spiegati con parole semplici e concetti pratici, come cerco di fare sempre io, in modo che siano di facile comprensione anche ai neofiti. Avendo fatto studi genetici in università avrei potuto usare anche io concetti più complessi ma certamente di difficile comprensione, non son voluto scendere su come ci comportano i cromosomi il DNA ecc, o piazzare complesse tavole di mendel senza alcuna spiegazione pratica. Questo articolo è nato proprio da una grande esigenza dei neofiti, che mi riempiono di domande sulla selezione e genetica dei Betta , perchè purtroppo appunto gli articoli su altri siti non li capiscono, e spesso in giro sui forum leggo cose fuori dal mondo.
Uno fra i tanti che sempre mi pongono domande di genetica, è l’ utente del nostro forum Simone a cui dedico questo articolo, dato che di domande me ne fa a sacchi . Se qualcosa in questo articolo non è chiara o qualcuno dice che ho espresso qualche concetto sbagliato lo invito a parlarne sul nostro forum , tutto quello che ho detto è frutto di mie esperienze nella selezione, incrociando le varie forme. Mi auguro che il l'articolo sia chiaro e utile a tutti.
Prossimo passo sarà un articolo sulla genetica delle colorazioni, ma credetemi li è ancora più complesso il discorso perchè le colorazioni sono tante in diverse combinazioni.
ATTENZIONE!!! E' SEVERAMENTE VIETATA LA COPIA ANCHE PARZIALE DEL TESTO, DEI DISEGNI E DELLE FOTO IN CUI NON E' RIPORTATA UNA FONTE ESTERNA A QUESTO SITO, IN QUANTO E' TUTTO DI PROPRIETA' DI BettaItalia.it . Se volete pubblicare questo o altri articoli sul vostro sito, blog o rivista contattateci: Questo indirizzo email è protetto dagli spambots. È necessario abilitare JavaScript per vederlo.





